|
|
CIVE 633 - ENVIRONMENTAL HYDROLOGY
NUTRIENT CYCLES
|
- Phosphorous is the most limiting nutrient in fresh water.
- The annual productivity of fresh waters is controlled more by P than any other nutrient or environmental factor.
- P can be stored in the sediments, and released slowly later.
- P reactions at the sediment-water interphase are important to understand the phosphorous cycle and budget.
- P is separated into several forms in the analytical process primarily through mechanical filtration.
- The orthophosphate anion (PO4-3) is the form available for uptake.
- The result from the undigested filtrate fraction is referred to as "soluble reactive phosphorous" (SRP)
and is analogous to dissolved inorganic phosphorous (DIP).
|
SRP --> DIP (DOP is usually small)
|
- TP ranges from 5 mg/L (5 ppm) in sewage effluent (most of which is SRP) to as little as 0.005 mg/L (5 ppb) in remote oligotrophic lakes.
- A detection level of at least 0.002 mg/L (2 ppb) is mandatory in lake research.
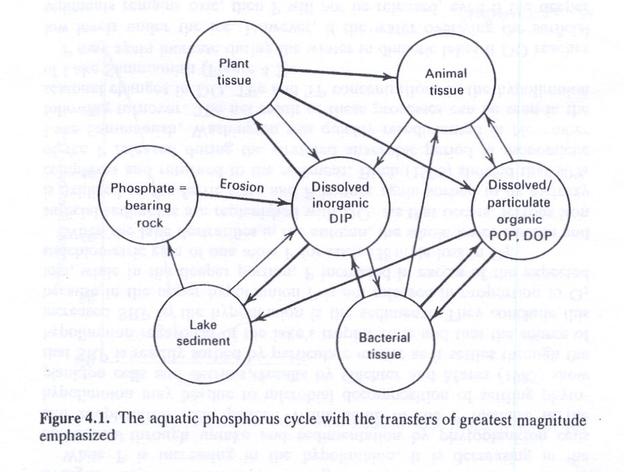
- Fig. 4.1 shows the aquatic phosphorous cycle.
- The ultimate source of P to aquatic ecosystems is from phosphate rock.
- P is utilized through plant uptake of DIP associated with photosynthesis, chemosynthesis, and decomposition.
- All organisms require P for metabolism and structure.
- Photosynthesis is mostly responsible for the uptake of DIP.
- Macrophytes and bacteria can also remove DIP from the water.
- Phytoplankton and bacteria are consumed by animal grazers, who in turn are consumed by predators.
- A fraction of DIP can enter the organic pools (DOP, POP) though excretion and death.
- DOP and POP can be recycled to DIP by bacteria.
- PIP and POP can settle into the sediments, and eventually converted to DIP.
- DIP can be released from the sediments.
- Sedimentation of dead phytoplankton and fecal pellets from phytoplankton and zooplankton result in loss of P from the open water.
- Physico-chemical conditions at the sediment-water interphase determine the release of P back to the water column.
|
4.1.3 SEDIMENT-WATER INTERFACE PROCESSES
|
- The exchange of P between sediment and water depends on several factors, acting separately or together:
- redox potential
- pH
- water exchange, as it affects diffusion and transport
- temperature, as it affects microbial activity
- relative fractions of P in the sediment that are bound with iron/aluminum, calcium and organic matter.
- The relative importance of these factors varies with depth and degree of thermal stratification.
- At the onset of thermal stratification, dissolved oxygen (DO) declines in the hypolimnion due to microbial decomposition of organic matter.
- As DO approaches 0, reducing conditions prevail and iron in the surficial sediments is reduced from its ferric form (Fe3+)
to is ferrous form (Fe2+).
- P, that was bound to the hydroxy complexes of ferric iron is now solubilized and released into the intersticial pore water, and is available for
diffusion into the overlying, anoxic water.
- The rate of diffusion is a function of the concentration gradient in SRP between the intersticial pore water and the overlying water.
- The hypolimnetic P content increases more or less linearly throughout the stratified period.
- Rates of release attributed to the iron redox process are variable.
- Values as high as 52 mg/m2/day with mean of 16 mg/m2/day have been reported.
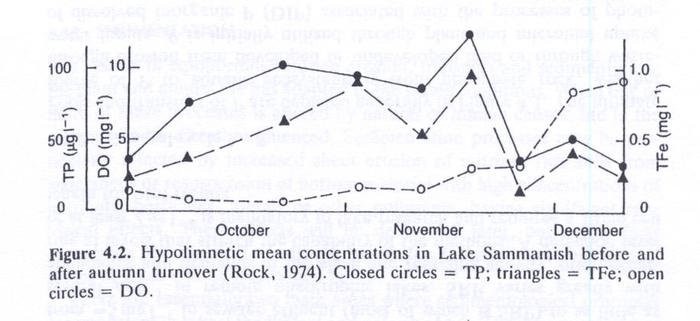
- When the lake destratifies in the autumn, the whole water column and surficial sediments are replenished with DO.
- Ferrous iron is then oxidized to the ferric state and P is once again sorbed in the hydroxy complexes and returned to the sediments (Fig. 4.2).
- Under oxic conditions, the solubility of iron is controlled by pH.
- At pH= 6, the solubility of ferric iron is minimal, and P can be effectively removed from the water column.
- With increasing pH, the solubility of iron increases and P is released from the sediments.
- High photosynthetic rates in eutrophic lakes can increase pH to 10, which produces high rates of release of P from the sediments.
- High pH due to photosynthesis can maintain high P in the water column.
- Temperature can be important in the release of P from sediments.
- The role of temperature is related to the stimulation of bacterial activity.
- Large release of P from sediments during summer can be attributed to iron-redox.
- In Lake Trummen in Sweden, removal of 1-m of rich sediment quickly resulted in recovery of the lake.
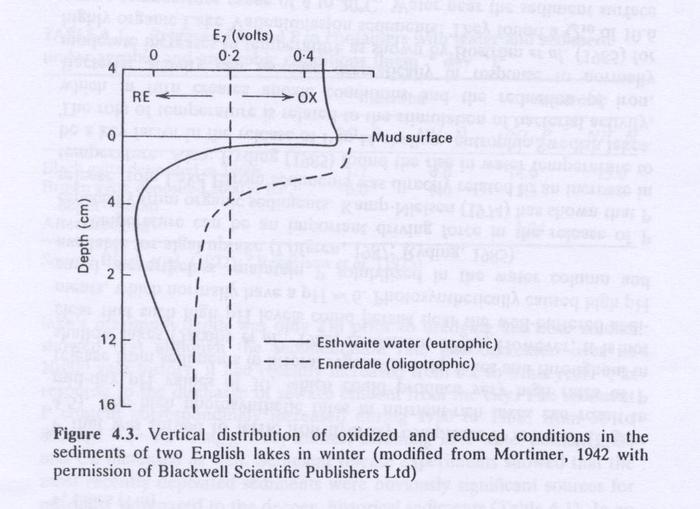
- Fig. 4.3 shows the redox potential in sediments and overlying water in two English lakes.
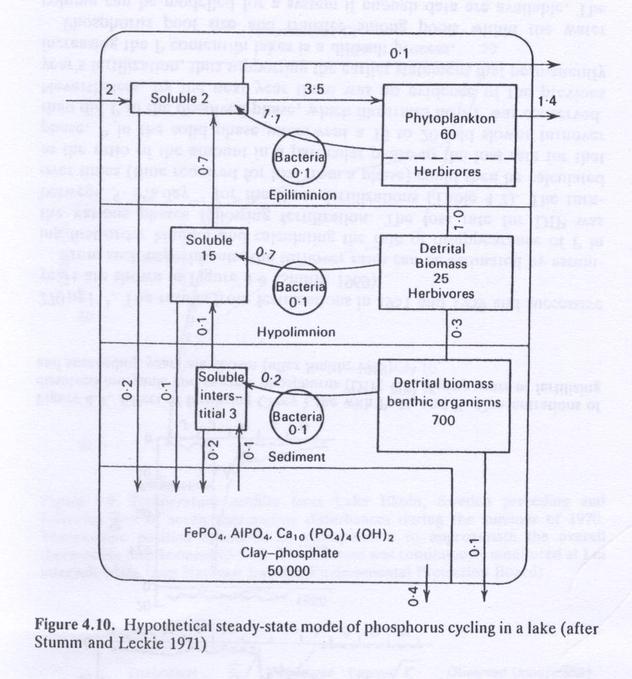
- Fig. 4.10 shows a hypothetical steady-state model of phosphorous cycling in a lake.
- Conclusions regarding sediment as source or sink of P:
- Sediments are nearly always a sink for P.
- Sediments can act as significant sources during a portion of the year.
- Whether the sediment-released P actually reaches the photic zone and is available for algal uptake is a significant issue.
- So long as sediments reach the hypolimnion, they are technically a source of P.
- There is ample evidence to support the statement that some sediment-released P is transported to the epilimnion.
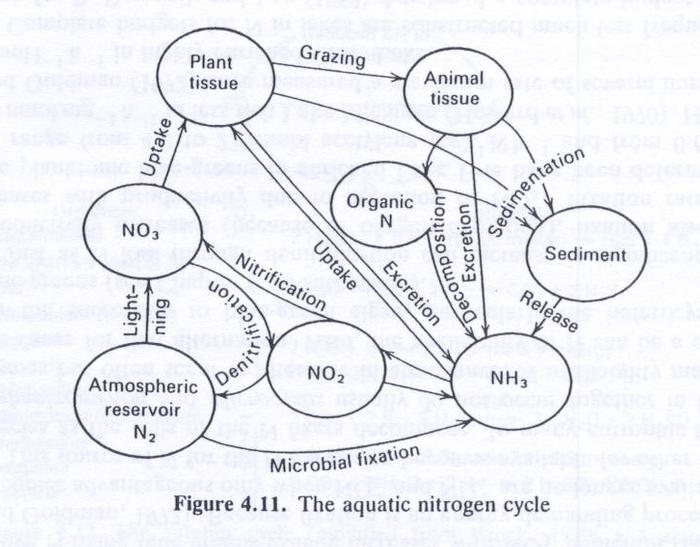
- The nitrogen cycle (Fig. 4.11) is very complex.
- Two large biological source and sink processes occur with N that do not occur with P.
- There are:
- The microbial fixation of N2 from the atmosphere.
- The return of nitrogen to the atmosphere through denitrification.
- The processes of nitrification (oxidation of ammonia) and denitrification may occur without biological mediation but at a slow rate.
- Microorganisms greatly speed up these processes.
- Nitrogen is most abundant as N2, constituting 78% of the atmospheric gases.
- Nitrate (NO3-) is the form of nitrogen that can be used by plants.
- Its concentration varies from a trace when productivity is high, to 1 mg/L when not used.
- Concentrations above 1 mg/L are usually associated with artificial inputs (fertilization).
- Ammonia (NH3) or ammonium (NH4+), which is the principal form in water,
becomes abundant in the absence of DO or in very enriched waters, but it is usually less abundant than nitrate.
- Plants often prefer ammonium to nitrate because it is in a more reduced form.
Nitrification
- Nitrification is the process by which NH3 is transformed first into NO2 and then into NO3.
- The process occurs only under aerobic conditions.
- Organisms that normally perform the transformations are Nitrosomonas and Nitrobacter.
- The yield of energy by nitrification is rather low compared to other transformations in the nitrogen cycle.
|
2NH4+ + 3O2 --> 2NO2- + 2H2O + 4H+ + energy
|
|
2NO2- + O2 --> 2NO3- + energy
|
Denitrification
- Denitrification occurs only in the absence, or near absence, of oxygen.
- A common denitrifier is Thiobacillus denitrificans.
|
5S2- + 6NO3- + 2H2O --> 5SO42- + 3N2 + 4H+ + energy
|
- In denitrification, the bacteria reduce nitrate first to nitrite and then to molecular nitrogen or N2O (nitrous oxide).
- This process removes nitrogen from ecosystems or wastewater.
- The necessary alternation of aerobiosis for nitrification, and anaerobiosis for denitrification has implications for management.
- Example: The Llanos de Mojos "camellones".
Nitrogen fixation
- Nitrogen fixation is an energy-consuming aerobic process carried on in aquatic environments by bacteria such as Azotobacter and Clostridium
and by blue-green algae (Cyanobacteria) Nostoc and others.
- Nitrogen fixation can represent a significant input of N to an ecosystem.
- Measured rates are from 0.00004 to 0.072 mg/L/day.
- In Clear Lake, in California, nitrogen fixation contributed 43% of the annual N input.
- Because N fixation is an energy-demanding process, it becomes advantageous only when nitrate or ammonium are no longer available.
- N fixation also increases with productivity as nitrate is depleted.
Implications to nutrient limitation
- In a freshwater lake, the residence time of N tends to be longer than P, because P tends to be removed to the sediments.
- N, as nitrate or ammonium, is much more soluble than P.
- N is limited in enriched waters through denitrification, but not as much in waters of low to moderate enrichment.
- N fixation from the atmosphere occurs in aerobic environments.
- N fixation occurs only when ecosystem is depleted of alternate sources of N.
- N occurs in precipitation more than P.
- In phosphorous-poor watersheds, little amounts of P in rain can be important.
- Nitrate in rain is very common, having been transformed from N2 to NO3- in the atmosphere by lighting.
- There are fewer sources of P than for N, and sedimentation is probably a more efficient remover of P than N in most aquatic ecosystems.
- In moderately enriched systems, P should be limiting.
- In highly enriched systems, the recycling of P and the loss of N through denitrification contribute to N being limiting.
- S is almost never a limiting nutrient in aquatic ecosystems.
- The normal levels as SO42- are more than adequate to meet plant needs.
- The sulphur cycle is shown in Fig. 4.12.
- Odorous conditions are readily created when waters are loaded with organic waste to the point that DO is removed.
- Then SO42- is the electron acceptor used for the breakdown of organic matter.
- H2S is produced, which has the smell of rotten eggs.
- If nitrate is available, N-reducing bacteria will dominate and odors will be minimal.
- Production of SO42- does not persist in the presence of oxygen.
- SO42- enters aquatic ecosystems through atmospheric deposition of sea salt and as a combustion product of fossil fuels,
manifested as acid rain, as well as through natural weathering processes in the watershed.
- Carbon (C) comprises nearly 50% of the dry organic matter in living organisms.
- C is usually not limiting to growth.
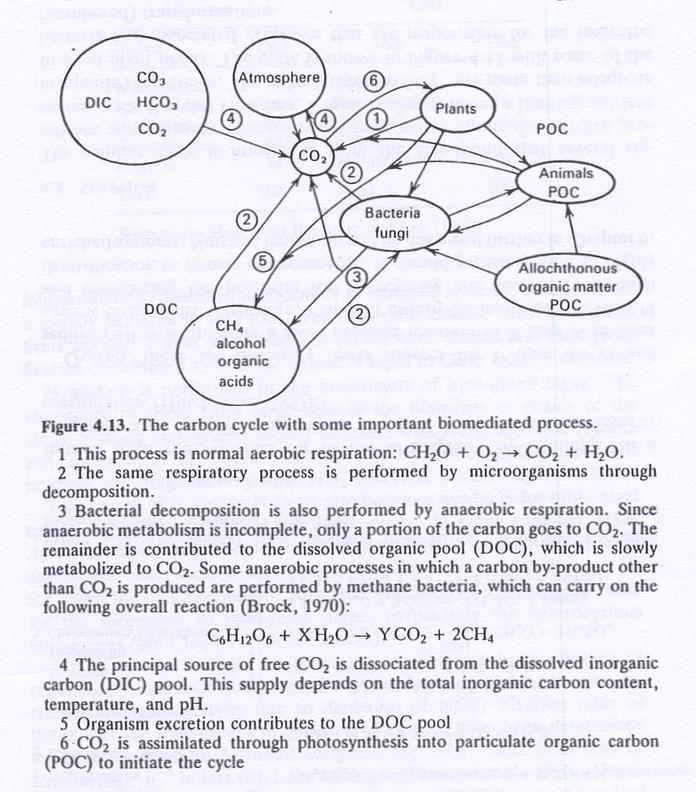
- Fig. 4.13 shows the carbon cycle.
- The atmosphere is a source of C, as with N.
- Rate of input of CO2 is dependent on the physical process of diffusion across the air-water interface.
- CO2 currently (2010) comprises 0.04% (400 ppm) of the atmospheric gases.
- Carbon is present in aquatic ecosystems as CO2, HCO3- and CO32-.
- CO2 diffuses into water from the atmosphere when the water is undersaturated, and from water to atmosphere when supersaturated.
- In eutrophic lakes, CO2 is depleted to very low levels causing pH to rise to 10 or more.
- Photosynthesis and respiration are two major factors that cause a significant departure from equilibrium of the system with the atmosphere.
- As algae phosynthesize, depleted CO2 can be replaced in water by the following two reactions:
- If CO2 is replenished from the atmosphere as fast as it is removed by algae,
the pH will not change.
- If algae consume CO2 faster than it can be replaced by diffusion from the atmosphere, H+ will decrease and
the pH will rise. Algal uptake usually does exceed atmospheric resupply.
- The pH will decrease if CO2 production through respiration is in excess of the CO2 loss through diffusion to the atmosphere.
- Photosynthesis tends to be self-limiting; if CO2 is reduced, the pH rises leading to more reduction of CO2.
|